|
『キュウリの芽生えの重力形態形成(CsPIN1)』編
これまでの地上実験により、キュウリの芽生えが重力に反応してペグを作る位置を決めるとき、オーキシンを輸送するタンパク質であるPINタンパク質のうち、”CsPIN1”と呼ばれるタンパク質が、ペグ形成時のオーキシン輸送に関わる可能性が考えられました。
なぜなら、種子を横向きにおいて発芽させて根と胚軸の境界域の下側にペグをひとつつくる条件にすると、CsPIN1タンパク質は上側で、より多く蓄積するようになるからです。
幼根が真下にでるように種子を縦向きにして発芽させた場合は、そのようなCsPIN1タンパク質の発現の違いはみられません。
この重力によって制御されるCsPIN1タンパク質の局在がオーキシンの再分布を誘導して、ペグ形成にかかわる可能性があります(図1)。
しかし、実際に重力によって、CsPIN1タンパク質の蓄積が変わるかは、宇宙で実験してみないとわかりません。
そこでCsPINs宇宙実験では、キュウリの芽生えのペグ形成部位におけるCsPIN1タンパク質の蓄積に対する重力の影響を検証し、重力による形態形成のネガティブコントロールの仕組みを理解することが目的です。
|
|
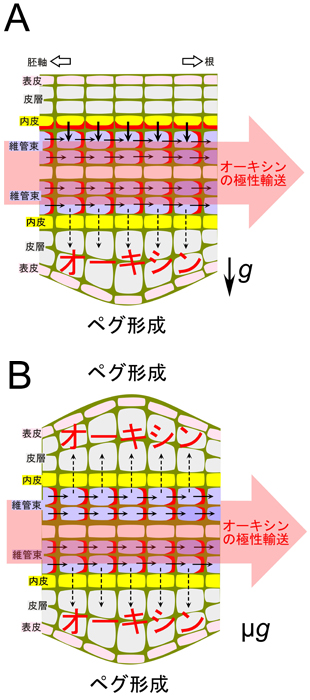
図1 CsPIN1タンパク質の重力応答性により制御されるオーキシン分布に関するモデル
キュウリ芽生えの境界域では、胚軸側から根側へと維管束を通って、オーキシンは極性輸送される。
地上で種子を水平において発芽させた芽生えの根と胚軸の境界域の上側において、重力刺激を感受する内皮細胞の維管束側の細胞膜にCsPIN1タンパク質が局在し、維管束から皮層・表皮へのオーキシンの流れを止める。
これにより、境界域の上側の皮層・表皮ではオーキシン量が増加せずに、ペグ形成が抑制されると考えられる。
一方、地上で種子を水平において発芽させた芽生えの根と胚軸の境界域の下側、あるいは微小重力環境下で発芽させた芽生えの根と胚軸の境界域の内皮細胞では、顕著なCsPIN1タンパク質の蓄積は起こらず、維管束から皮層・表皮へのオーキシンの流れが積極的に抑えられない。
その結果、境界域の皮層・表皮ではオーキシン量が増加し、ペグ形成が誘導されると考えられる。
|
|
|
『キュウリの芽生えの根の水分屈性(CsPIN5)』編
根が曲がるのは、根の両側で伸び方が違うために起こります。
つまり、一方で伸び方が大きく、一方で小さくなるために、その結果として曲がります。
これを偏差成長といい、オーキシンの不均等な分布が原因となって誘導されます。
根ではオーキシンのより多い側の成長が相対的に小さくなって、オーキシンの多い側に曲がるようになります。
背景で述べたように、横向きにおかれた根では、オーキシンが伸長域の下側に多く輸送されるようになり、水分勾配下の根では、オーキシンが伸長域の高水分側に多く輸送されるようになると考えられます(図2)。
したがって、地球上で重力屈性と水分屈性が干渉し合う現象は、重力と水分勾配によって制御されるオーキシン動態の競合によると考えられます。
そこで、これらの現象を理解するためには、重力と水分勾配がどのような仕組みでオーキシンを下側、高水分側に輸送するのかを理解することが重要になってきます。
実は、この決まった方向へのオーキシンの輸送にも、オーキシン排出担体が関係しています。キュウリ芽生えの根で、オーキシン排出担体の細胞膜への蓄積を解析したところ、CsPIN5タンパク質が、根の先端から伸長域の表皮に蓄積し、それがオーキシンを根の先端から伸長域に運ぶように局在することがわかりました。
興味深いことに、このCsPIN5タンパク質の蓄積は、横向きにおかれた根では伸長域の上側で減少し、水分勾配下では低水分側で減少する可能性が示されました。
これらの結果は、横向きの根の下側、水分勾配下での高水分側でオーキシン量が相対的に多くなることに、このCsPIN5タンパク質の関与することを示唆しています。
そこでCsPINs宇宙実験では、上記の仮説を検証するために、微小重力下でキュウリの芽生えの根の重力屈性と水分屈性を分離し、CsPIN5タンパク質の蓄積に対する重力刺激と水分勾配刺激の影響を明らかにすることが目的です。
|
|
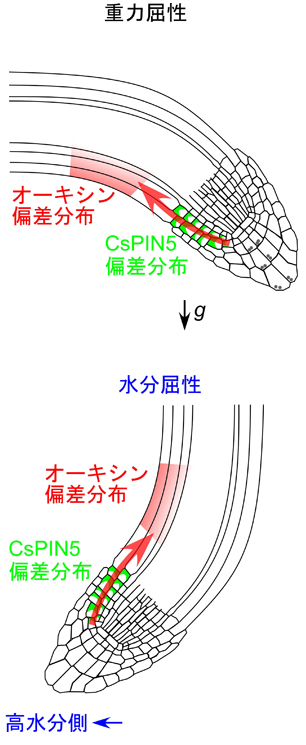
図2 環境刺激に応答したオーキシン偏差分布の形成に関するモデル
重力刺激、あるいは湿度勾配に応答して、より多く水分を含む側でCsPIN5が多く蓄積し、より多くのオーキシンが輸送される。
この結果、オーキシンの偏差分布が形成され、根が屈曲する。
|
|



